
Research Article | DOI: https://doi.org/10.31579/2637-8876/018
1 Department of laboratory Medicine, HangzhouMedical College, Hangzhou,310053, China.
2 Centre of Laboratory Medicine,People’s Hospital of Hangzhou Medical College, Zhejiang Provincial People’s Hospital,
*Corresponding Author: Dazhi Jin, Department of laboratory Medicine, Hangzhou Medical College. No. 481 Binwen Rd, Hangzhou, Zhejiang, 310053, China.
Citation: Shuangshuang Wan, Guangzhong Song, Hui Hu, Yaqing Xu, Peng Zeng, et al. (2021). Intestine Epithelial cell-Derived Extracellular Vesicles alleviate Inflammation induced by Clostridioides difficile TcdB THROUGH the Activity of TGF- β1, J. Immunology and Inflammation Diseases Therapy. 4(1); Doi:10.31579/2637-8876/018
Copyright: © 2021 Dazhi Jin. This is an open-access article distributed under the terms of The Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Received: 11 October 2021 | Accepted: 11 November 2021 | Published: 17 November 2021
Keywords: extracellular vesicles; Clostridioides difficile; TGF- β1; TcdB; regulatory T cells; inflammatory cytokines; immunotherapy 36
Background: Clostridioides difficile infection (CDI) has been primarily associated with the toxin B (TcdB), which can activate the intestinal immune system and lead to pathological damage. Even though the biological functions of intestine epithelial cell- derived extracellular vesicles (I-Evs) have been well documented, the role of I-Evs in the process of CDI is still unknown.
Results: We isolated I-Evs ranging from 100–200 nm in mean diameter, with a density of 1.09-1.17 g/mL. These I-Evs expressed the extracellular vesicle-associated specific surface markers, CD63 and TSG101. In vitro, 50 µg I-Evs decreased the expression of IL-6, TNF- β, IL-1β, and IL-22 in MC38 induced by 0.8 ng/mL C. difficile TcdB, and increased expression of TGF- β1. In vivo, I-Evs also promoted regulatory T cell induction, which improved inflammation of mice up to 80% relative to C. difficile TcdB infected mice, depending on the TGF- β1 signal pathway.
Conclusion: Our study firstly demonstrated that I-Evs originated from intestine epithelial cells is potentially a novel treatment endogenous candidate to effectively reduce the local infection induced by C. difficile TcdB.
In recent decades, with the excessive application of broad-spectrum antibiotics, diseases related to intestinal flora disorders have precipitously increased. Clostridioides difficile (C. difficile) is one of the main pathogens leading to antibiotic-associated diarrhoea and hospital-acquired infections in the United States and other developed countries [1]. Toxin A (TcdA) and B (TcdB) are the major pathogenic factors leading to diarrhoea, pseudomembranous colitis, toxic megacolon, and other intestinal symptoms [2]. The mechanism lies in the inactivation in the host epithelial cells of proteins from the Rho family of GTPases-including Rho, Rac, or Cdc42 by glycosylation, and upregulation of a series of pro-inflammatory cytokines such as interleukin IL-1, IL-6, and TNF-α [3]. Meanwhile, toxins recruit neutrophils and other inflammatory immune cells to induce intestinal mucosal cell apoptosis, necrosis, shedding, and increased permeability, triggering a widespread loss of intestinal barrier function, and initiating imbalance of flora and intestinal epithelial damage. According to the American Infectious Society, and the European Society of Clinical Microbiological Infections, in addition to other practical guidelines, oral metronidazole or vancomycin are the best methods to treat Clostridioides difficile infection (CDI) [4]. In addition, some new narrow-spectrum antibiotics such as fidaxomicin [5] and rifaximin have little impact on the intestinal flora and reduce the risk of drug resistance. In recent years, a number of immune-based agents [6] have entered clinical trials, and however their efficacy needs to be further validated. Faecal microbiota transplantation (FMT) has been recognised in the United States as an optional treatment method to restore normal intestinal flora and prevent recurrent attacks. However, a meta-analysis of randomised clinical trials in 2019 showed that the cure rate of FMT was only 76.1%. Furthermore, there are still many unanswered questions about FMT, including the optimal timing, preparation methods, and the patients who are likely to benefit most from this procedure. As its standard protocol is relatively complicated and involves approval of ethical reviews, FMT has not yet been widely used in China. Extracellular vesicles (Evs) are small vesicle-like substances secreted by cells, which possess various biological activities when released outside of the cell. They have a diameter ranging from approximately 30 nm to1 µm, and are generally classified into exosomes, microvesicles, and apoptotic bodies based on their size, biogenesis, and mechanism of secretion [7]. It is difficult to determine the functional differences between these three types of Evs, due to the lack of specific markers with which to distinguish them. Although once thought to be cellular debris, Evs are now recognized as vital vehicles involved in the communication between cells. Research has confirmed that Evs contain a wide range of biologically active components, and their corresponding functions depend on the source tissue or cell type. Evs also exist in body fluids such as serum, alveolar lavage fluid, and breast milk, carry messenger RNAs, microRNAs, and DNA [8, 9]; this suggests potential applications as biomarkers for the diagnosis of diseases, as part of a liquid biopsy technology [10]. Recently, it has been reported that Evs can be designed to function as effective carriers in the treatment of various diseases, including in the delivery of long non-coding RNAs [11, 12]. In addition, Evs play a significant therapeutic role in regulating complex intracellular pathways in certain diseases, such as inflammatory bowel disease (IBD) [13, 14], and osteoarthritis [15]. Furthermore, it has been discovered that Evs derived from mesenchymal stem cells possess important immunomodulatory effects in areas such as neurodegenerative diseases, ageing, and inflammation [16-18]. Previously, we have reported that CD8α+CD11c+ Evs derived from lungs reduce the allergic reaction of asthmatic mice through TGF-β1 and IL-10, thereby maintaining the immune balance of the respiratory tract [19]. In the context of the recent outbreaks of COVID-19 around the world, mesenchymal stem cells and their Evs could be used as potential drug candidates for the treatment of severe cases, mainly through the induction of anti-inflammatory macrophages, regulatory T and B cells, and regulatory dendritic cells [20]. Strikingly, infection with TcdB-producing strains alone, but not TcdA+B- strains, can cause severe CDI symptoms [21]. Our work using purified C. difficile TcdB, together with cell lines and mice, confirmed that TcdB can induce expression of the inflammatory genes IL-6, TNF- β, IL-22, and IL-1 β, and upregulation of TGF-1 in vitro. Intestine epithelial cell-derived extracellular vesicles (I-Evs) rescue this phenomenon in vivo by inducing proliferation of regulatory T cells, dependent on TGF- β1 and the corresponding downstream molecules Smad2/3. Here, we studied the role of I-Evs on inflammation induced by C. difficile TcdB and evaluated biological functions of I-Evs in alleviating pathological damage led by CDI in mice.
Results
Isolation and identification of intestine epithelial cell-derived extracellular vesicles We used electron microscopy to visualise the morphology of the purified I-Evs; combined with Nanoparticle tracking analysis this showed that the isolated I-Evs had a mean diameter of 100–200 nm (Figure 1A, B). To further explore the I-Evs, sucrose density gradient centrifugation was used to detect the density range of I-Evs, which was 1.09–1.17 g/mL (Figure 1C). Protein analysis identified that I-Evs were positive for universal surface markers of extracellular vesicles, including CD63 and TSG101, and the intestinal epithelial cell-specific protein A33, but negative for GRP94, as detected by western blot (Figure 1D).
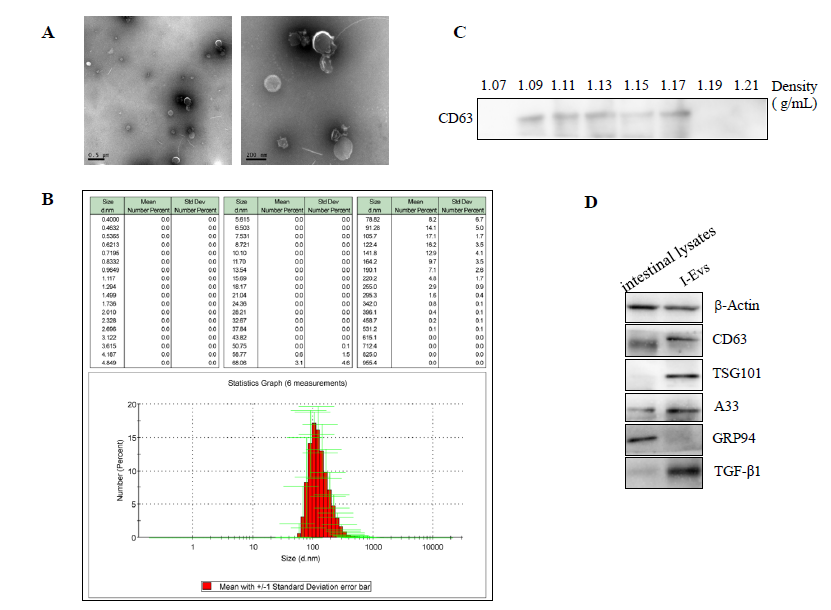
In addition, high levels of TGF-β1 were expressed in I- Evs, implying a role in immunoregulation. The results showed that we successfully isolated and identified I-Evs. I-Evs attenuated the down-regulation of TGF- β1 induced by purified C. difficile TcdB in vitro Real-time PCR results showed that, compared to the control group, the expression of pro-inflammatory genes (IL-6, TNF- β, IL-1β, and IL-22) was increased in the 0.4 ng/mL C. difficile TcdB group, but significantly decreased in the 0.8 ng/mL I-Evs group. In contrast, the expression of the anti-inflammatory genes TGF- β1 and IL-10 was statistically increased in the I-Evs group compared to the TcdB groups (Figure 2A).
Western blot results showed that protein levels of the immunosuppressive cytokine TGF- β1 were decreased in MC38 murine colon carcinoma cells, and LOVO human colon carcinoma cells, stimulated by C. difficile TcdB (Figure 2B). The concentration of C. difficile TcdB used was 0.1 ng/mL, 0.2 ng/mL, 0.4 ng/mL, or 0.8 ng/mL. This decrease could be rescued by I-Evs when TcdB concentration was 0.4 ng/mL (Figure 2C). Altogether, these results indicate that the I-Evs containing TGF- β1 had anti- Inf ammatory effects in vitro.

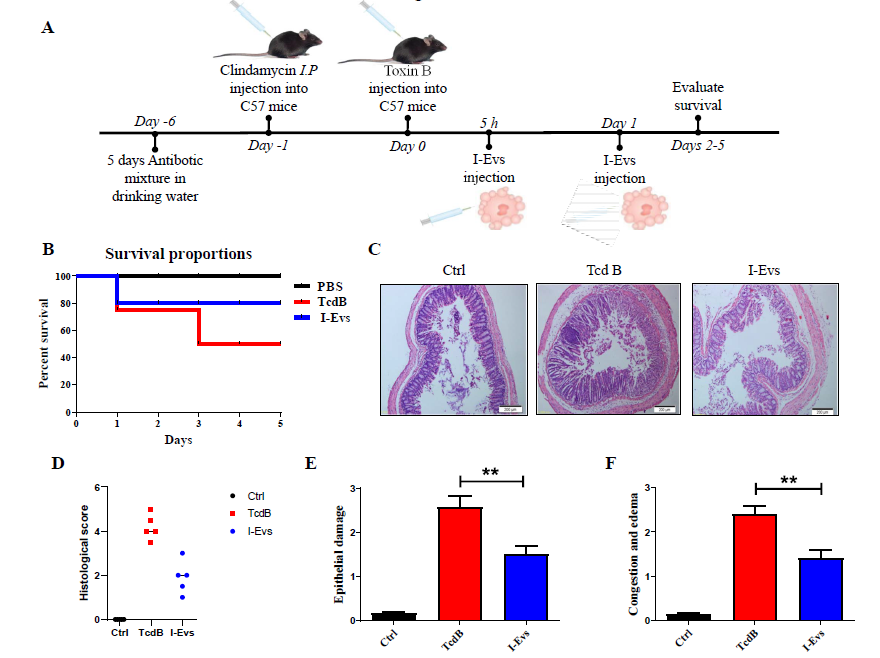
I-Evs alleviate C. difficile TcdB-induced local colon inflammation in mice
Intestinal epithelial damage caused by C. difficile TcdB, generally confined to the local intestine, is a severe inflammatory intestinal lesion. We sought to explore whether I-Evs can be applied in this condition as a type of anti-inflammatory immunotherapy. I-Evs contain more TGF- β1 than intestinal lysates as determined by western blot, which indicated a likely strong immunosuppressive effect. Next, we established a murine local colon infection model to investigate the treatment effect of I-Evs (Figure 3A). As shown in Figure 3B, the survival rate of mice after C. difficile TcdB injection was only 50%, while I-Evs increased the survival rate of mice up to 80%. The intestinal tissues displayed marked leukocyte infiltration and sections of glandular structure damage; consistently, histopathological analysis showed only slight leukocyte infiltration and epithelial cell damage after application of I-Evs (Figure 3C, D). Moreover, intestinal epithelial damage, congestion and mucosal oedema were significantly increased in the C. difficile TcdB mice when compared with the control mice (Figure 3E, F), however, less intestinal damage and limited leukocyte infiltration were observed when mice were treated with I-Evs. These findings implied that I-Evs attenuated pathological changes occurring as a result of C. difficile TcdB-induced inflammation, thereby protecting mice from local colon inflammation.
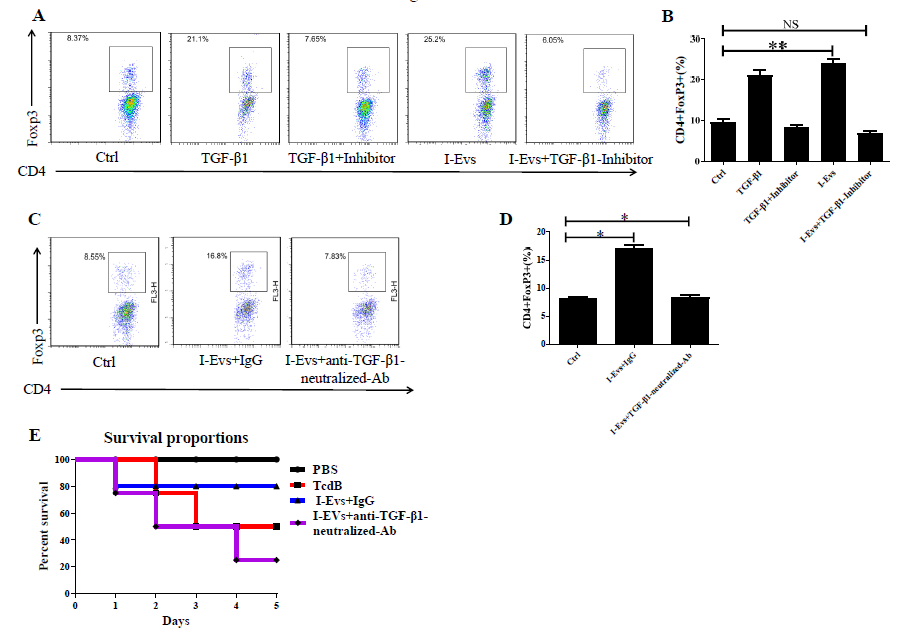
Induction of regulatory T cells by I-Evs alleviated infection caused by C. difficile TcdB through a TGF-β1-dependent mechanism
A previous study showed that EpCAM-dependent I-Evs alleviated IBD by inducing regulatory T cells [22]. I-Evs induced an increase in the proportion of CD4+Foxp3+Tregs in vitro and in vivo (Figure 4A–D); these immunoregulatory cells exhibit immunosuppressive effects in the development of disease. When the activity of TGF-β1, a potent immunosuppressive cytokine, was blocked (using the protocol described in the Materials and Methods), I-Evs immediately lost the ability to induce CD4+Foxp3+Tregs in the spleen. Concurrently, I-Evs were not able to increase the survival rate of mice, and the improvement of pathological effects previously seen was also undetectable (Figure 4E, F,4G).
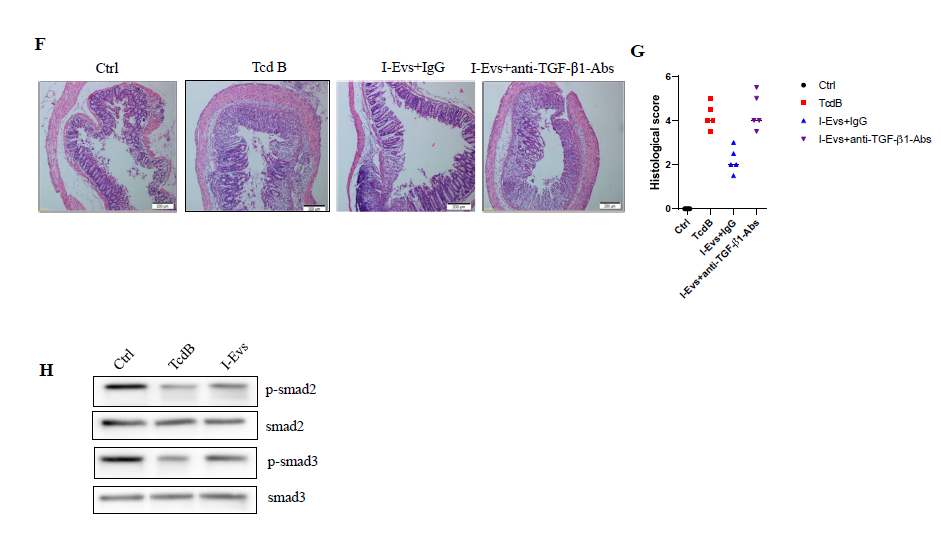
Together, these results suggest that immunosuppressive regulatory T cells induced by I-Evs attenuated C. difficile TcdB- induced local colon inflammation in a mechanism dependent on TGF-β1. Smad2/3 are the main downstream proteins involved in the TGF- β 1 signalling pathway. The phosphorylation levels of Smad2/3 were decreased after stimulation with C. difficile TcdB, although protein levels of Smad2/3 remained the same; treatment with I-Evs promoted phosphorylation of Smad2/3, and thereby upregulation of TGF- β1 (Figure 4H). These results suggest that Smad2/3 is inhibited by C. difficile TcdB, leading to the down-regulation of TGF- β1 expression. Conversely, I-Evs with high expression of TGF- β1 activate Smad2/3 and contribute to the upregulation of TGF- β1, there by alleviating C. difficile TcdB-induced local colon inflammation in mice.
A common clinical symptom of CDI is local colon infection, which may arise due to intestinal perforation after either infection or intestinal surgery, particularly in high- risk populations, such as patients with IBD; respiratory insufficiency; heart and renal failure; ages over 60 years; and several other underlying diseases. The majority of CDI can be treated with metronidazole and fidaxomicin, in addition to other antibiotics. Surgical removal of necrotic intestinal tissue can reduce mortality rates with severe explosive colitis. Nevertheless, postoperative bleeding, and intestinal stenosis and obstruction, are extremely distressing to the patient. Prevention, management, and non- surgical treatment are the fundamental principles of CDI. However, the most severe toxic colitis cases are unable to benefit from drugs and surgery, and there is an urgent need to establish an effective treatment programme based on immunotherapy. Evs participate in a variety of physiological and pathological processes, including neurological disorders [23], osteoarthritis [24], infection [25], and tumours [26]. Evs have been proven to be involved in immune regulation and antigen presentation, and our research group demonstrated that Evs derived from intestinal epithelial cells alleviate IBD in mice by inhibiting dendritic cell activation and inducing Tregs [22]. In this study, I-Evs isolated from the intestine, with mean diameters of 100–200 nm as detected by electron microscopy scanning and Nanoparticle tracking analysis, expressed the characteristic protein markers of Evs, CD63 and TSG101. Enrichment of the immunosuppressive cytokine, TGF- β 1, in I-Evs inspired us to hypothesise an immunomodulatory function for I-Evs. A recent study verified that Evs derived from human mesenchymal stem cells can relieve colitis by reducing pro-inflammatory responses and increasing anti-inflammatory responses [27]. As is well-established, colitis caused by C. difficile relies on a series of virulence factors, including toxins, which initially target intestinal epithelial cells and subsequently destroys the intestinal membrane integrity. Hosts exposed to intestinal microorganisms trigger immune inflammatory responses. The dominance of either TcdA or TcdB was still controversial in this research field, despite a multi-laboratory follow-up research study pronouncing that TcdB acts as a critical toxin in colonic epithelial injury and mortality in vivo, whereas TcdA caused inflammation in mice to a small extent [28]. In the work presented here, TcdB induced increased gene expression of IL-6, TNF- β, IL-1β, and IL-22. I-Evs were able to rescue this phenomenon, and interestingly, TGF- β 1 and IL- 10 gene actually increased upon co-incubation with I-Evs. Moreover, I-Evs could reverse the decreased expression of TGF-β1 stimulated by C. difficile TcdB, as detected by western blot analysis of MC38 and LOVO lysates. We also report for the first time that I-Evs can improve survival of mice with local colon inflammation induced by C. difficile TcdB. The mechanism lies in the induction of CD4+Foxp3+Tregs, which play an important role in maintaining immune tolerance and homeostasis; the decline or dysfunction of Tregs has previously been shown to increase intestinal inflammation in IBD mice[29]29. Similarly, CD4+CD25+ Treg cells transferred into hosts ameliorated colitis symptoms. The I-Evs in this study contained TGF- β1; IL-10 is also known to be an important immunosuppressive cytokine, but we could barely detect the presence of IL-10 in our isolated I-Evs. Whether IL-10 still performs an important function is unknown. Furthermore, proinflammatory cytokines were undetectable following stimulation with C. difficile TcdB. Indeed, we improved various experimental schemes to optimise the experimental conditions, unfortunately, the corresponding data were still not available. We speculate that the effect of C. difficile TcdB on cells in vitro was different from that in vivo. As for the animal challenge experiment, in order to induce chronic inflammatory intestinal infection, five antibiotic mixtures were fed to mice, in addition to intraperitoneal injection with clindamycin and local injection with C. difficile Tcd B. I-Evs improved both the survival of mice, and intestinal tissue pathological scores, when transferred into mice.
We firstly demonstrated that I-Evs can alleviate inflammation induced by C. difficile TcdB in vitro and in vivo, and protect against pathological lesions in the animal intestine. Our data indicated that I-Evs could activate the TGF- β1 pathway and the downstream proteins Smad 2/3, to alleviate local colon inflammation induced by C. difficile TcdB, providing a novel endogenous candidate for treatment of C. difficile infections.
Toxins, antibodies, and reagents
C. difficile TcdB was gifted from the Tao Liang research group (West Lake University, Hangzhou, China) [30]. Primary antibodies against CD63 (ab213090), TGF-1 (ab8227), GRP94 (ab238126), TSG101 (ab125011), and β-Actin (ab8227) were from Abcam (Cambridge, MA, USA). PRMT1 (A33) (#2449), Smad2 (D43B4), Smad3 (C67H9), phospho-Smad2 (Ser465/Ser467) (E8F3R), and phospho-Smad3 (Ser423/425) were from Cell Signalling Technology (Danvers, MA, USA), and the corresponding secondary antibodies were from BBI (Shanghai, China). Fluorescent- labelled antibodies against CD4 (GK1.5) and Foxp3 (PCH101) were from eBioscienc (San Diego, CA, USA).
Real-time fluorescence quantification PCR
The classic TRIzol (Gibco, USA) method was used to extract RNA, using a reverse transcription kit (TOYOBO) to acquire cDNA. Real-time, fluorescence quantification PCR (qRT-PCR) was performed in a Step One Plus Real Time PCR System (Roche) to detect gene expression. The mouse-specific primers used are shown in Appendix A.
Mouse and cell lines
The MC38 cell line was purchased from Wuhan Fine Biotech Co., Ltd. (Wuhan, China). The cells were negative for mycoplasma as detected by fluorescence and culture methods. Human LOVO cells were kindly provided by Jia Jing (Hangzhou Medical College, Hangzhou, China). Male C57BL/6J mice (6–8 weeks old) were purchased from Shanghai Laboratory Animal Co., Ltd. (Shanghai, China). The mice were housed in a specific pathogen-free animal facility, and experimental protocols were approved by the Animal Care and Use Committee of Hangzhou Medical College, all animals were treated according to the guidelines for animal experimentation of Hangzhou Medical College in Hangzhou, China. The animal experiments were also performed in accordance with the ARRIVE (Animal Research: Reporting of In Vivo Experiments) guidelines [31].The mice were sacrificed 5 days after anesthetized with intraperitoneal injection chloral hydrate (375 mg/kg of body weight)
Isolation and quantification of I-Evs
Mouse large intestines were surgically extracted and ground in a sufficient volume of PBS. They were then digested with 1 mg/mL collagenase type Ⅱ from Clostridium histolyticum (Gibco) for 2 h at 37 ˚C. The resulting suspension of intestinal tissue fragments was centrifuged at 400 g for 10 min, and the supernatant carefully removed for further centrifugation at 10,000 g for 30 min, to remove larger vesicles. The resulting supernatant was then filtered by a 0.22-µm screening and ultracentrifuged at 100,000 g for 1 h. Crude pellets of I-Evs were washed in sterile PBS and centrifuged at the same speed for an additional 1 h. The harvested I-Evs were resuspended in PBS. A BCA assay was used to detect the concentration of I-Evs (ThermoFisher, Waltham, MA, USA).
Electron microscopy scanning and Nanoparticle tracking analysis
Suspensions of I-Evs were loaded onto a coated copper grid, and a drop of 2% phosphotungstic acid added as a negative staining method. The sample was then allowed to dry at room temperature and transferred to a transmission electron microscope (Hitachi H7650, Hitachi, China) to take pictures and record at a voltage of 80 kV. To detect size distribution, I-Evs were diluted with PBS, and 0.3 mL analysed by NanoSight Nano instruments (Malvern, UK).
Western blot and flow cytometry analysis
For western blot analysis, 40 µg I -Evs or protein lysates extracted from intestinal tissues were separated by 12% sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE), and transferred to a polyvinylidene fluoride (PVDF) membrane (Millipore, Billerica, MA, USA). Membranes were blocked with 5% milk in phosphate buffered solution-Tween 20 (PBS-T) and incubated with the corresponding primary antibodies at 4 ˚C overnight. The next day, membranes were incubated with an HRP-coupled secondary antibody for 1 h at room temperature and scanned using a Canon 4500 imaging system (Shanghai, China). For cytometry analysis, cells were washed with cold PBS and incubated with a fluorescent antibody for 30 min at 4 ˚C in the dark. Cells were analysed by fluorescence-activated cell sorting (BD,Franklin Lakes, NJ, USA).
CD4+Foxp3+Tregs induction assay
Murine CD4+ T cells were isolated with the EasySep Mouse CD4+ T Cell Isolation Kit (Stemcell), and labelled with an anti-CD62L antibody for flow cytometry. Magnetic sorting was then performed using the EasySep Mouse Biotin Positive Selection Kit (Stemcell). Cells were then incubated with 1 µl anti -CD3/CD28-coated beads and 200U/mL IL-2 for 72 h (2 x 105 cells/well), with or without 50 µg/mL I -Evs. To block the TGF- β1 signal, 0.6 µg/mL TGF- β1 inhibitor was applied to cells (in vitro), or 15 µg/mL anti–TGF- β1–neutralising antibody was injected into mice (in vivo). The percentage of CD4+Foxp3+Tregs was analysed by flow cytometry.
Induction and treatment of murine local colon inflammation induced by C. difficile Tcd B
C57BL/6J male mice were randomised into groups and given antibiotics through their drinking water for 5 days. The antibiotic mixture consisted of gentamicin (0.035 mg/mL), kanamycin (0.4 mg/mL), colistin (850 U/mL), metronidazole (0.215 mg/mL), and vancomycin (0.045 mg/mL) (Sigma-Aldrich, St. Louis, MO, USA). The following day, mice were injected with clindamycin (10 mg/kg). After this, purified TcdB was surgical injected into local colon of mice (0.5 µg/kg); this was noted as day 0. Functional I-Evs (50 µg/100 µL PBS) were intraperitoneal injection after 5 h, and on day 1. After sacrificing the animals, the intestinal tissue in different groups was collected and prepared for H&E staining.
Data are presented as the mean ± SEM. Data were compared using a Student’s t- test with GraphPad Prism 8 (San Diego, CA, USA). P<0.05 was considered statistically significant.
C. difficile: Clostridioides difficile; CDI: Clostridioides difficile infection; FMT: Faecal microbiota transplantation; Evs: Extracellular vesicles

Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
